Lesson 13.3: The Origin of Species
Lesson 13.3: The Origin of Species
Lesson Objectives
Recognize that new discoveries since Darwin have added an understanding of speciation to evolutionary theory.
Explain the concept of a species.
Define the biological species concept and analyze its usefulness.
Compare the biological species concept to morphological, genealogical, and ecological concepts.
Analyze the reasons why biologists consider all humans to be members of the same species.
Define speciation.
Describe two conditions that can lead to speciation.
Explain the results of speciation in terms of adaptation, chance, and changes in the environment.
Distinguish allopatric from sympatric speciation.
Describe an experiment which demonstrated allopatric speciation.
Describe two general types of reproductive isolation.
Explain how polyploidy can result in sympatric speciation.
Discuss the use of hybridization to form new crop species.
Analyze the importance of environmental complexity to sympatric speciation for ani- mals.
Compare and contrast the gradualist and punctuated equilibrium models of evolution- ary change.
Describe conditions that could increase the rate of speciation.
Describe circumstances that could lower the rate of speciation.
Species Concept
Darwin avoided the use of the word evolution in his major work about evolution. The word “evolve” appears once, at the very end. He titled his work The Origin of Species – a process or group of processes now called speciation.
What exactly is a species? How did the millions of species which exist on earth today arise? Will they (we!) continue to change? How quickly? These are questions Darwin sought to answer – and the questions we will explore in this lesson on evolution. Darwin’s work opened the door to life’s history. This lesson will look at the details and modifications of his ideas which research has made clear in the 150 years since he published The Origin.
What is a Species?
A toddler readily recognizes that she/he is surrounded by distinct groups of individuals which we call “kinds” of plants and animals. Biologists refer to kinds of living things as a species - groups of individual organisms which are very similar.
If we look closely, however, variations make it quite difficult to decide where to draw the lines between species. For example, are a St. Bernard and a Chihuahua similar enough to group within the same species? Just how similar must two organisms be for biologists to decide they are members of the same species? The answer to this question is not clear, even to biologists who specialize in classification. We will explore several, because how we define a species can help to clarify how species develop.
The Biological Species Concept
A widely used definition of species is the biological species concept (Figure 13.28), first proposed by evolutionary biologist Ernst Mayr in 1942. Mayr’s concept begins with the idea that members of a species must be similar enough that they could interbreed. Because all dogs
from a St. Bernard to a Chihuahua – are capable of interbreeding, biologists consider all dogs to be members of the same species. If you are familiar with mules, however, you know
that this definition needs clarification. If horses and donkeys mate, they produce mules, but mules are sterile and cannot continue to interbreed. Therefore, the biological species concept becomes organisms similar enough to interbreed and produce fertile offspring. Horses and donkeys, therefore, are not members of the same species. As you may know, wolves and dogs can interbreed to produce viable hybrids with fertile offspring; surely, wolves and dogs are not members of the same species? The last part of the definition addresses this problem, and the complete definition becomes:
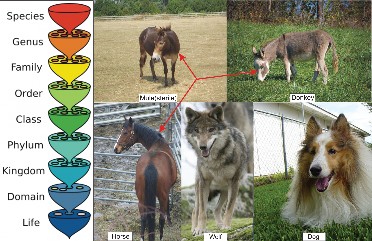
A biological species is a group of organisms similar enough that they could interbreed and produce fertile offspring under natural conditions.
This definition serves the goal of defining members of a species as individuals which are still undergoing evolution – they form a distinct yet potentially common gene pool. You will learn much more about classification in the next chapter, but here it is important to realize that one of the primary goals of classification is to show evolutionary history – patterns of common ancestry. This makes the biological species – a functionally reproducing unit – an important foundation of classification. Closely related species descended from relatively recent common ancestors, and distantly related species descended from more distant common ancestors. The emphasis the biological species concept places on successful reproduction fits this goal of classification quite well.
Another advantage of the biological concept is that it has the potential to explain how speciation occurred – that is, how two closely related groups of organisms became different species through reproductive isolation. Members of two species may appear very similar, yet fail to interbreed because of reproductive barriers. Barriers to reproduction may either prevent mating, or prevent development of a fertilized egg after mating. Elaborate courtship behaviors, including the blinking pattern of fireflies or the songs of birds, are often required to elicit mating behavior – and limit mating to members of a species (Figure 13.29). Different breeding seasons, such as flowering dates, can also prevent interbreeding. Molecular differences between even closely related species may prevent sperm or pollen from actually fertilizing eggs. Once the eggs are fertilized, chromosomes may be so incompatible that mitosis and meiosis cannot proceed normally; if the zygote cannot develop, offspring do not survive. All of these barriers between species work to ensure successful reproduction within species, keeping specific, useful adaptations “in the family,” so they are a logical way for us, too, to distinguish members of one species from members of another.

Although the biological species concept is extremely helpful in evolutionary thinking it has serious limitations for practical use. For organisms that reproduce asexually – including all bacteria and viruses - the definition is entirely unworkable. Nor can we detect whether or not fossil organisms would have been able to interbreed – whether or not they coexisted. Biologists must rely on structure and (if available) biochemical similarities to classify fossils and most microorganisms.
The Morphological Species Concept
Alternatives to the biological species concept emphasize the characteristics and processes which unite, rather than divide (reproductive barriers), species. We will look at just a few in order to gain insight into evolutionary thought. A much more practical definition is the morphological species concept, which groups organisms based on structural and biochemical similarities. Recent advances in molecular biology, such as DNA comparison, have strengthened this means of clarifying evolutionary relationships. Biologists probably use this method more than any other to differentiate species in nature, despite its limitations
in confirming the potential for interbreeding.
The Ecological Species Concept
The ecological species concept focuses on a group’s common ecological niche – the set of environmental conditions and resources used or required by the group. This concept is based on the idea that ecological and evolutionary processes divide resources in such a way that individuals can most efficiently adapt to use those resources as a group. All members of a species, then, have a unique set of adaptations to a particular set of environmental conditions. Note that both the morphological and ecological definitions “work” for asexually reproducing organisms, and many fossils, as well. However, they do not help to explain how two closely related groups became different species, as does the biological definition.
The Evolutionary Species Concept
The last concept we will consider has the potential to clarify the path of speciation, or evo- lutionary history – that primary goal of classification. The genealogical or evolutionary species concept defines a species as a group of organisms with a unique genetic history
a group which shares common ancestry without divergence. A species, according to this concept, is a group which forms one tip on the branching tree of life’s history. Modern technology which compares DNA and protein sequences makes this definition workable, but still, the mechanism of speciation is not defined.
Ideally, all of these ways of determining exactly which individuals make up a species would merge; all make valid points about the idea of a species, and all seek the common goal of defining a new, unique unit of life in space and time. Practically, however, each has its own usefulness for different purposes. If we were working in the field, we would undoubtedly use the morphological concept. However, as we explore the idea of speciation mentally and through lab models, we will adopt the biological species concept as our working definition.

What about humans? Are we all members of the same species? According to the biological species concept, the answer is a resounding yes. Despite our differences in appearance, culture, and location on planet Earth, any human female is capable of producing fertile offspring with any human male. Therefore, all humans are members of the same biological species (Figure 13.30). In contrast, humans and chimpanzees do not interbreed even when they share the same territory, so biologists consider chimpanzees to be a distinct species. Before we leave the “species problem” and Homo sapiens, it is worth noting that all of the various definitions of species confirm the unity of the human species. DNA sequences, cell chemistry, anatomy, ecological resources, and reproductive ability all reveal similarities which unite us as a single species with common ancestry. In fact, the most recent and precise method of comparison – DNA sequences – shows that somewhere between 1 in 100 and 1 in 1,000 base pairs differ, on average, between one human and the next. Based on this analysis, we humans are 99.9% identical. The differences which seem so great to us (skin color, facial features, build, personality) are important for recognition of individuals, and a few may adapt us to particular regions of the Earth, but overall, we are vastly more similar to than different from each other; we are one species.
Although we may not learn exactly how we came to be a separate species, we can gain some insight by looking closely at what is known of the process of speciation. What do we know now – that Darwin did not know - about “the origin of species”?
An Overview of Speciation
Speciation occurs at the boundary between microevolution and macroevolution. At what point do the allele frequency changes of microevolution add up to define a new and separate species?
Ernst Mayr, who developed the biological species concept mentioned above, also identified two essential components of speciation. For two populations to evolve into separate species, they must experience:
Isolation limits gene flow which would tend to maintain uniformity between the two pop- ulations. Physical separation by a barrier limits migration and prevents interbreeding, but differing environmental conditions may also provide a form of isolation. Isolation increases the chance that two populations will take separate microevolutionary pathways. Returning to our white and brown rabbits, even a difference in altitude and thus temperature might separate two parts of a population so that albino individuals will be favored higher on a
Isolation
Genetic divergence
mountain where snow frequently accumulates, but brown individuals would be favored down toward the valleys, where snow rarely covers the ground.
Although the two rabbit populations might be isolated by environmental differences, they would not become separate species unless enough genetic differences accumulate to pre- vent successful interbreeding. Genetic divergence is more likely if populations are small, increasing the chance of genetic drift. Even in large populations, however, environmental differences may result in large genetic differences which prevent interbreeding. In our rabbit populations, for example, selection for white rabbits might be accompanied by mate selec- tion based on white coat color. If mate preferences were heritable and sufficiently strong, interbreeding between the brown population and the white population would cease, and the two populations would become two species.
The result of speciation is two groups of individuals, each more closely adapted to local environmental conditions than the larger parent population. Although speciation may not always result in a better fit between a species and its environment (remember that genetic drift increases the influence of chance), over time, natural selection tends to increase fitness. A species’ adaptations to both physical and living environments increase the chance that the species will survive. The millions of species which exist today are each carefully attuned to a particular niche. And, just as variation within a species ensures that some individuals will survive environmental change, a great diversity of species increases the chance that at least some organisms will survive major changes in the environment.
Isolation and genetic divergence are both prerequisites for speciation, but they are so closely related that sometimes it is difficult to distinguish one from the other. Physical isolation may encourage genetic divergence, but many evolutionary biologists believe that reproductive isolation can result from genetic divergence alone. That leads us to two major categories of speciation – one based on geographic isolation, and the other based on reproductive isolation alone. These two categories, and examples of speciation from each, are the topic of the next section.
Isolating Mechanisms
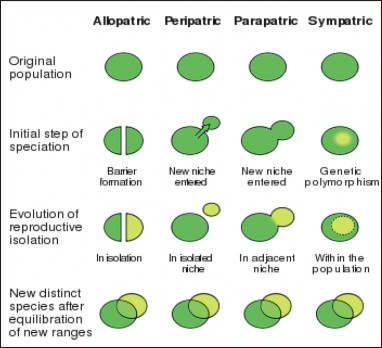
Biologists today divide isolating mechanisms into two major categories based on whether they happen in different locations (allopatric = “other fatherland”) or a single location (sympatric
= “same fatherland”) (Figure 13.31).
Allopatric Speciation
Allopatric speciation involves geographic barriers, which physically isolate populations. Formation of a land bridge such as the Isthmus of Panama, for example, could separate members of a marine population into two groups. Emergence of a mountain range could separate members of a lowland species. According to the fossil record, the rifting of continents
divided populations of terrestrial animals and plants. In these cases, dramatic changes in landforms lead to geographic isolation; however, movements of animals and plants can also result in physical isolation. Single individuals or small groups of individuals may move away from a parent population to colonize a new area, and the new colony could be isolated from its parent population. Such movements are not always intentional; a storm apparently carried a small flock of finches from the coast of South America to the Galapagos Islands. In each case, geographic barriers (the land bridge, the ocean) isolate the two populations so that gene flow stops and genetic divergence can proceed. Natural selection may lead each population to adapt to its own unique environment, or genetic drift may lead to chance differences in gene pools. If genetic divergence results in reproductive incompatibility, the two populations have become two separate species.
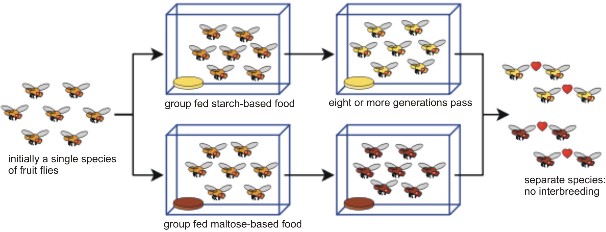
Diane Dodd, working with a laboratory population of fruit flies (Figure 13.32), experimen- tally verified the idea that physical isolation can lead to reproductive isolation and speciation. Dodd split the population into two groups, and fed maltose to one group and starch to the other. She observed that each new environment selected for improved efficiency in digesting the available food molecule. After eight or more generations of isolation, the two populations were recombined. After isolation, “starch” flies preferred to mate with other “starch” flies, and “maltose” flies preferred “maltose” mates. Although the preferences were not absolute, they did demonstrate at least initial formation of a reproductive barrier.
Sympatric speciation involves the emergence of a new species within the geographic range of the parent population. In the absence of geographic isolation, reproductive barriers must arise in different ways in order for new species to form. Some biologists doubt the relative im- portance of sympatric speciation to evolutionary change, but several examples demonstrate the potential for this mechanism of evolution.
Polyploidy
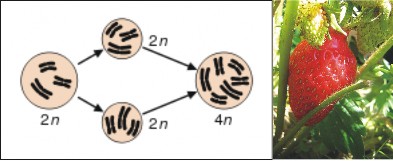
In plants, new species occasionally arise by duplication of chromosomes – a condition known as polyploidy (Figure 13.33). Recall that our human body cells are diploid, and that egg and sperm cells are haploid. If meiosis fails to reduce the number of chromosomes, diploid sex cells result. In plants, which can self-pollinate, diploid pollen may fertilize a diploid egg, resulting in a tetraploid offspring. Although tetraploids may self-pollinate or interbreed with other tetraploids, they cannot successfully reproduce with their parents, because three sets of chromosomes (two from the tetraploid parent and one from the “normal” diploid parent) cannot successfully perform the intricate dance of meiosis. Peanuts, potatoes, and cotton are familiar crops that are tetraploid. Strawberries and sugarcane are octaploid (eight sets of chromosomes in each cell, compared to our two)! Polyploidy is a common form of speciation in plants in nature, as well as in agriculture.
Two species of plants may hybridize – often forming unusually vigorous hybrid offspring. Unfortunately, despite their vigor, these offspring are often infertile due to chromosome incompatibility. However, if polyploidy occurs within the offspring, each of the previously unmatched chromosomes has a partner, offspring fertility is restored, and a new species is formed. Triticale (Figure 13.34), a hexaploid hybrid of wheat and rye, was produced in the laboratory in this manner. Combining the high yield and quality of wheat with the disease- resistance of rye, triticale is now a successful grain crop in Europe, China, and Australia.

Polyploidy is less common in animals, perhaps because they less frequently self-fertilize. Some salamanders are polyploid, but offspring are usually sterile and “species” reproduce by parthenogenesis (development from unfertilized eggs). Human and other mammalian livers often contain many polyploid cells.
Although polyploidy is rare among animals, other zoological modes of sympatric speciation exist. For example, environmental change within a population’s range may lead to two habitats in which genetic divergence may take place. Our hypothetical rabbit example in the last section is an example of this type of sympatric speciation. A field example involves the hawthorn fly (Figure 13.35), whose larval maggot stage feeds on hawthorn fruit. Apple trees, introduced to the US, often grow near hawthorns, and some hawthorn flies have begun to feed on apples. The two populations, although sympatric, have developed significant genetic differences, and at least in the wild, no longer tend to interbreed. This apparent reproductive isolation involves both temporal divergence (apples and hawthorns and their respective fly populations mature in slightly different seasons) and mating and egg-laying preferences (which link larval fruit to adult behavior). Many biologists consider these flies to be an example of sympatric speciation in progress.
Cichlid fish in East Africa’s Lake Victoria illustrate both allopatric and sympatric speciation (Figure 13.36). Less than a million years old, the Lake harbors nearly 200 closely related cichlid species. Biologists conclude that adaptive radiation by a small group of colonizers

into available niches explains the diversity of feeding specializations among these closely related species, much like Darwin’s finches in the Galapagos. Because the various habitats in the huge Lake (or islands in the oceans) isolate populations before speciation, adaptive radiation is often considered a type of allopatric speciation. However, nonrandom mating may have led to at least one more recent sympatric speciation. Females of two closely related species appear to select mates based on differing, brightly colored backs. Although the two color variations can still interbreed in the lab, they do not appear to do so in nature.

The Tempo of Speciation
Speciation and extinction characterize all life on earth; the fossil record clearly documents both. Two startling facts emerge from careful study of the fossil record: First, the average successful species lives for “just” a few million years. Second, over 99.9% of all species that have ever lived have become extinct. The last aspects of speciation that we will consider are the tempo and pattern of species formation.
Over time, geographic changes isolate populations. Small populations experience genetic drift. Mutations alter individual genotypes and gene pools. New habitats form, and small groups colonize them. It is clear that evolution continues to change life. However, there is considerable debate about the rate at which speciation occurs over geologic time. Most biologists agree that single mutations seldom if ever cause new species in single evolutionary “leaps.” Mutations in regulatory genes, which have major effects during development, may be an exception, but in general, mutations are more likely to be harmful, and selected against. Except for the special case of polyploidy, discussed above, speciation cannot occur within a single generation. So, what do we know about the rate and pattern of speciation?
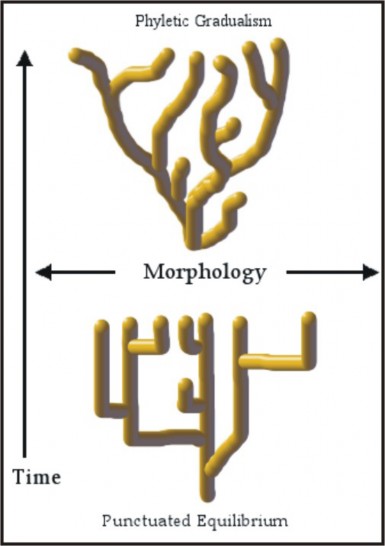
Some evolutionary biologists consider the rate of evolution to be slow and constant, with small changes accumulating to form big changes – the idea of gradualism. Others (Niles Eldridge and Stephen Jay Gould), in response to the apparently “sudden” appearance of new forms in the fossil record, suggest that species diverge in bursts of relatively rapid change, and then remain stable for relatively long periods – a model known as punctuated equilibrium. Gould maintains that speciation and evolution occur rapidly in small, peripheral populations, whereas large, central populations remain stabilized for long periods of time. It is the large, central, stable populations which are represented in our fossil record, he argues – not the small, peripheral, evolving ones. The two models are compared in Figure 13.37.
In either case, species require several thousand generations – which may require several thousand years – to form. Relative to the species’ “lifespan” and the modest precision of our ability to pinpoint time within the geologic record, the differences between the two models may be minimal. Major characteristics of species could have evolved gradually over thousands of years, and still we would not be able to detect those changes on the geologic time scale. What we do detect are long periods of relative stability, when stabilizing selection tends to maintain equilibrium. Changes in physiology and internal anatomy could have continued during the long periods of apparent equilibrium without our seeing them; the fossil record cannot reveal these types of changes. The debate is not about the mechanism of evolution
only its tempo.
Proponents of either model admit that the pace of evolution probably changes from one set of conditions to another. Lake Victoria in East Africa, for example, is just a million years old, and yet home to more than 300 species of cichlid fish found nowhere else (return to Figure 13.35). Genetic analysis shows that all of them descended from a single ancestral “founder” species – relatively rapid evolution. Without competition, that species expanded across the huge Lake, and natural selection suited individual populations to exploit the many different food sources and habitats present in the Lake. Cichlid evolution is an example adaptive radiation similar to that of Darwin’s finches and Hawaiian honeycreepers, discussed in the Evolutionary Theory chapter. The fossil record shows similar opportunities for accelerated evolution after mass extinctions. The extinction of the dinosaurs opened an abundance of ecological niches, soon occupied by radiating birds and mammals. Speciation may accelerate not only with a major increase in available niches, but also with the appearance of new adaptation, such as legs or wings, which allows a group to enter a variety of new niches. In contrast, long periods of environmental stability may slow the pace of speciation.
If you look back at the opening questions for this lesson, you can see that both evolution and our investigations into how evolution works are ongoing - works in progress. We have come a long way in understanding “the origin of species” since Darwin, but questions remain. Exactly what is selected - genes, genotypes, phenotypes, or individuals – is still debated;
some, such as Stephen Jay Gould, accept that selection acts at multiple levels. Exactly how fast speciation occurs also remains under discussion; both gradualism and punctuated equilibrium models have merit. Several definitions of a species today serve different purposes; eventually, they should merge into a single clear picture of this basic unit of evolution.
Behind these disagreements, however, lies a powerfully reinforced theory of evolution and an increasingly detailed story of life on Earth. By any definition, humans form a single species, increasingly united by mobility, common genes, and common resources. We and the millions of species with whom we share the Earth today all arose from a single common ancestor billions of years ago – through deep time and myriad instances of allopatric and sympatric speciation. Amazing adaptive logic was most often a result of: directional, stabilizing, and disruptive environmental forces selected for favorable variations and against unfavorable ones. Chance played a major role – through mutation, gene flow, genetic drift, and much environmental change. The pace of speciation probably varied – from gradual change, to rapid change, to long periods of stability. Both speciation and extinction continue today – and will continue into the future, although we cannot predict the direction, because chance determines so much of the direction evolution takes. As a unique species, we are significantly influencing the amount of extinction – and may consequently affect the rate of speciation, as well. That, however, is a topic for a later chapter.
Lesson Summary
Ever since Darwin’s publication of The Origin of Species, biologists have continued to work out the details of how species originate – the foundation of evolutionary process.
A species is the smallest group of organisms into which biologists classify living things.
A biological species is a group of organisms that can interbreed to produce fertile offspring under natural conditions. This is probably the most widely accepted definition of a species.
A morphological species groups organisms based on extensive structural and biochem- ical similarities. This is probably the most practical definition for use in the field.
A genealogical or evolutionary species includes organisms which share a recent, unique common ancestor. This is the goal of all classification; the only question is how to recognize members.
An ecological species groups organisms together if they share a unique set of adapta- tions to a particular set of environmental conditions.
Ideally, all four definitions would merge to recognize a true species.
All humans are members of the same biological species, because all races and cultures can and do intermarry, and our DNA is 99.9% identical.
For populations to become separate species, they must experience isolation and genetic divergence.
Isolation can be physical or environmental, but it must be accompanied by heritable reproductive isolation, which requires genetic divergence.
The result of speciation is usually two groups of individuals, each more closely adapted to local environmental conditions than the larger parent population.
Because genetic drift and environmental change increase the influence of chance, adap- tation is seldom “perfect” – but over time, natural selection tends to increase fitness.
Allopatric speciation involves geographic barriers, which physically isolate populations.
Barriers to mating and barriers to development of zygotes can both cause reproductive isolation.
Experiments with fruit flies support the possibility that geographic isolation can lead to reproductive isolation.
Sympatric speciation involves the emergence of a new species within the geographic range of the parent population.
In plants, polyploidy is a common sympatric form of “instant speciation.”
Hybridization together with polyploid has formed many vigorous crop species.
In animals, environmental complexity may lead to sympatric speciation.
Cichlid fish in Lake Victoria illustrate both adaptive radiation – a form of allopatric speciation – and more recent sympatric speciation due to heritable changes in mating preference.
The Gradualism model of speciation holds that small changes accumulate to form big changes.
The Punctuated Equilibrium model suggests that rates of change accelerate over short periods in small peripheral populations, and then stabilize for long periods in large, central populations.
Differences between the two may be minimal, because gradual change can occur over thousands of years without our ability to detect it in the fossil record.
Conditions that may accelerate speciation are mass extinctions and the formation of new habitat (e.g. volcanic islands) because both create a sudden availability of many “empty” niches.
Evolution of a major new adaptation, such as legs or wings, may also accelerate speci- ation by suiting a species to multiple new habitats.
Long periods of environmental stability may slow the rate of speciation.
Review Questions
Further Reading / Supplemental Links
Dawkins, Richard. The Ancestor’s Tale: A Pilgrimage to the Dawn of Life. 2004. Boston: Houghton-Mifflin, ISBN 0618005838 Punctuated Equilibrium
Define the biological species concept and analyze its usefulness.
Compare the biological species concept to morphological, genealogical, and ecological concepts.
Analyze the reasons why biologists consider all humans to be members of the same species.
Describe two conditions that can lead to speciation.
Explain the results of speciation in terms of adaptation, chance, and changes in the environment.
Distinguish allopatric from sympatric speciation.
Describe two general types of reproductive isolation.
Describe the use of hybridization and polyploidy to form new crop species.
Analyze the importance of environmental complexity to sympatric speciation for ani- mals.
Compare and contrast the gradualist and punctuated equilibrium models of evolution- ary change. Give examples which support each.
http://www.geocities.com/ginkgo100/pe.html
http://www.pbs.org/wgbh/evolution/library/03/5/l_035_01.html
Phylogenetic Tree
Eastern and Western Meadowlark information, with songs - Cornell Lab of Ornithology
http://www.birds.cornell.edu/AllAboutBirds/BirdGuide/Western_Meadowlark. html
http://www.birds.cornell.edu/AllAboutBirds/BirdGuide/Eastern_Meadowlark.
Adaptive Radiation of Cichlids
http://www.current-biology.com/content/article/fulltext?uid=PIIS0960982207017046
http://www.nature.com/hdy/journal/v99/n4/full/6801047a.html
Vocabulary
allopatric speciation The evolution of a new species from a closely related population isolated by geographic barriers.
biological species concept A group of organisms similar enough to interbreed and pro- duce fertile offspring under natural conditions.
ecological niche The set of environmental conditions and resources used or required by a species; the role a species plays in its ecosystem.
ecological species concept A group of organisms which share a unique set of adaptations to a particular set of environmental conditions.
evolutionary species concept See genealogical species concept.
genealogical (evolutionary) species concept A group of organisms which share a re- cent, unique common ancestor – common ancestry without divergence.
gradualism The idea that the tempo of evolution is slow and constant, with small changes accumulating to form big changes.
hybrid Offspring of cross-breeding between two different but closely related species.
morphological species concept A group of organisms which share extensive structural and biochemical similarities.
polyploidy The duplication of chromosome sets, often resulting in “instant speciation.”
punctuated equilibrium The idea that species diverge in bursts of relatively rapid change and then remain stable for relatively long periods.
reproductive barrier A condition which prevents mating or prevents the development of offspring.
reproductive isolation The separation of closely related populations by barriers to pro- ducing viable offspring.
speciation The process which results in new, separate and genetically distinct groups of organisms (species).
species See biological, ecological, genealogical, and morphological species concepts.
sympatric speciation The evolution of new species from closely related populations lo- cated in the same area.
Which definition of species – biological, morphological, ecological, or genealogical – do you prefer?
To what extent do you think stabilizing, directional, and disruptive selection affect humans today?
What effects might genetic engineering have on speciation?
Do you find the evidence for sympatric speciation (the more disputed of the two forms) convincing?
Are gradualist and punctuated equilibrium models mutually exclusive?
Why don’t disagreements about speciation threaten the theory of evolution by natural selection?
Image Sources
http://commons.wikimedia.org/wiki/Image:MRNA-interaction.png. Public Domain.
National Park Service. http://commons.wikimedia.org/wiki/Image:Wolf_pack_in_Yellowstone_NP.jpg. Public Domain US-NPS.
{kind=link}
http://www.flickr.com/photos/zenera/352770831/. Public Domain, CC-BY-SA.
BGA. http://www.flickr.com/photos/benimoto/603735193/. Creative Commons.
Joel Rotunda. http://www.flickr.com/photos/jrotunda85/192554288/. CC-BY-SA.
http://commons.wikimedia.org/wiki/File: Wheat,_rye,_triticale_montage.jpg. GNU-FDL, Public Domain.
Benny Mazur. http://www.flickr.com/photos/benimoto/603735193. CC-BY.
User Toony, GFDL. http://commons.wikimedia.org/wiki/Image:Selection.svg. CC-BY-SA-2.5, 2.0, 1.0.
{kind=link}
http://commons.wikimedia.org/wiki/Image:Types-of-mutation.png. CC-BY-2.5, Public Domain NIH.
{kind=link}
Ted E. http://commons.wikimedia.org/wiki/Image:Population_bottleneck.svg. GNU-FDL.
{kind=link}
http://en.wikipedia.org/wiki/Image:Punctuatedequilibrium.png. PD-User.
{kind=link}
Phrood from the Talking Glossary of Genetics.
http://commons.wikimedia.org/wiki/Image:Chromosom.svg. Public Domain.
{kind=link}
Renato Bsasutti. http://commons.wikimedia.org/wiki/Image:Map_of_skin_hue_equi3.png. GNU-FDL.
{kind=link}
Brenda. http://www.flickr.com/photos/jiggapotpie/328581516/. CC-BY-SA.
http://commons.wikimedia.org/wiki/Image: Kr%C3%B3liki_kalifornijskie_666.jpg. CC-BY-SA-2.5, 2.0, 1.0.
Jan Roletto. http://commons.wikimedia.org/wiki/Image:See_elefanten.jpg. Public Domain.
{kind=link}
http://en.wikipedia.org/wiki/Image:Selection_Chart.png. Public Domain.
{kind=link}
National Institutes of Health, Department of Health and Human Services. http://commons.wikimedia.org/wiki/Image:HIV_Daughter_Particles.jpg. Public Domain-USGOV-HHS-NIH.
{kind=link}
Maureen. http://www.flickr.com/photos/amerune/302221440/. CC-BY.
Ilmari Karonen. http://commons.wikimedia.org/wiki/Image:Speciation_modes.svg. Public Domain.
{kind=link}
http://commons.wikimedia.org/wiki/Image:MasaiMaraCheetahs01.jpg. Public Domain.
{kind=link}
Shelley Rainey. http://www.flickr.com/photos/85853969@N00/1466951276. CC-BY.
http://commons.wikimedia.org/wiki/Image:HIV-SIV-phylogenetic-tree.png. Public Domain.
{kind=link}
Geneviève Baujat and Martine Le Merrer Ellis-Van, Creveld Syndrome, Orphanet Journal of Rare Diseases, 2007. http://commons.wikimedia.org/wiki/Image:Polydactyly_ECS.jpg. CC-BY-2.0.
{kind=link}
BGA. http://www.flickr.com/photos/benimoto/603735193/. Creative Commons.
http://commons.wikimedia.org/wiki/Image:Portrait_of_Muslim_man.jpg http://www.flickr.com/photos/73416633@N00/391989140/ http://www.flickr.com/photos/exfordy/155874181/http://www.flickr.com/photos/marcalandavis/63268510/. CC-BY-2.0,
CC-BY-SA-2.0, CC-BY, CC-BY, CC-BY, CC-BY.
http://commons.wikimedia.org/wiki/Image: Drosophila_speciation_experiment.svg. Public Domain.
http://commons.wikimedia.org/wiki/File:Mucoviscidose.PNG. The creator of this work allows anyone to use it for any purpose including unrestricted redistribution, commercial use, and modification, GNU Copyright-Free.
{kind=link}
http://en.wikipedia.org/wiki/Image:EasternMeadowlark23.jpg. PD USFWS, PD US Gov.
{kind=link}
(30) http://www.flickr.com/photos/7193008@N02/449542472/ http://www.flickr.com/photos/wrex/277392736/ http://commons.wikimedia.org/wiki/File:Canis_Lupus_SignatusZOOM.jpg http://www.flickr.com/photos/grendelkhan/106098744/ http://www.flickr.com/photos/35034360660@N01/33791657. CC-BY-SA-2.5.
Kazulanth. http://commons.wikimedia.org/wiki/Image:Founder_effect.png. Public Domain.
{kind=link}
http://commons.wikimedia.org/wiki/Image:Hb-animation2.gif http://commons.wikimedia.org/wiki/Image:Redbloodcells.jpg http://commons.wikimedia.org/wiki/Image:Sickle_cell_hemoglobin.png http://commons.wikimedia.org/wiki/Image:Sicklecells.jpg.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CC-BY-SA-2.5,2.0,1.0, Public Domain.
http://commons.wikimedia.org/wiki/Image:Hiv-monotherapy-levels.png. GNU-FDL.
{kind=link}
http://commons.wikimedia.org/wiki/Image:Allele-frequency-fr.png. GNU-FDL.
http://en.wikipedia.org/wiki/Image:Sickle_cell_distribution.jpg. CC-BY-SA-2.5, 2.0, 1.0, CC-BY-SA-2.5, 2.0, 1.0.
{kind=link}
User Waugsberg, GFDL. http://commons.wikimedia.org/wiki/Image:Bienen_mit_Brut_1.jpg. CC-BY-SA-2.5, 2.0, 1.0.
{kind=link}
http://commons.wikimedia.org/wiki/Image:Crataegus-monogyna-frugt.JPG http://commons.wikimedia.org/wiki/Image:Jab%C5%82ko_-_owoc.JPG. (a) Public Domain (b) GNU-FDL (c) CC-BY-SA-2.5.
{kind=link}
{kind=link}
- Log in or register to post comments
- Email this page